Ядоносный апарат змій складається з ядопродуцирующих залоз, вивідних проток і ядопроводящих зубів. Самий складний і досконалий ядоносный апарат у гадюкових (Viperidae) і ямкоголовых (Crotalidae) змій. Зуби у них укріплені на короткій верхньощелепної кістки, здатні обертатися, при закриванні рота лягають вздовж щелепи вістрям назад, при розкритті піднімають і приймають прямовисне, «бойове» положення. Для отруйних ужеобразных змій (Colubridae) характерні борознисті зуби, розташовані на верхній щелепі ззаду; для аспидовых (Elapidae) і морських (Hydrophiidae) змій - короткі нерухомі трубчасті зуби в передній частині верхньої щелепи. Довжина зубів у звичайної гадюки 0,5 см, у габонській - 3-4 див. Вони тонкі, гострі і крихкі; легко, як голка, встромлюють в м'які тканини, при зустрічі з твердим об'єктом ковзають і ламаються. Зламані зуби, як і природно, що випали, замінюються розташованими поруч зубами-заступниками. Всередині або на передній поверхні зубів є канал. У гадюкових він розташований глибоко і відкривається у кінця зуба.
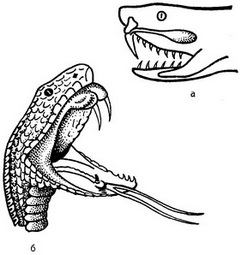 Рис. 9. Голова отруйної змії. а - видно залоза, що виробляє отруту; ядоносный канал, який відкривається в основу ядоносного зуба; б - відкрита паща гюрзи; видно два ядоносных зуба і мову. |
На підставі зубів знаходяться протоки ядопродуцирующих залоз, що складаються з численних бульбашок різних форми і розміру (рис. 9). У гюрзи, піщаної ефи, степової та звичайної гадюки ядопродуцирующая заліза розгалужена трубчаста, з одним вивідним протокою. У гадюк задній її відділ більше переднього. Каудальные трубочки переднього відділу утворюють перегородки типу клапанів, завдяки чому не відбувається змішування секрету відділів при спокійному стані. При гистохимическом дослідженні у відділах виявлена лужна фосфатаза, в сполучнотканинних перегородках - ацетилхолинэстераза; в секреті заднього відділу ацетилхолінестерази мало, а в передньому відділі її немає зовсім. У передньому відділі передбачається наявність гіалуронідази, є провідником нейротоксин через гематоенцефалічний бар'єр, про що свідчить відсутність токсичного секрету заднього відділу до змішування його з нетоксичним секретом переднього відділу (А. М. Захаров, 1968). Отже, токсичний ефект досягається шляхом змішування секретів, що відбувається при укусі.
Кількість отрути в залозах залежить від фізіологічного стану змії, часу знаходження її на волі і пройшов з моменту останнього укусу, від статі і віку змії, кліматичних умов і т. д. У голодних змій отрути більше, ніж у ситих, у самців більше, ніж у самок (р <0,05), при цьому токсичність майже однакова. Залишковий отрута значно менш токсичний (Wiener, 1960). В, залозах у звичайної гадюки отрути близько 30 мг, у піщаної гадюки і кобри - до 200 мг; у гримучої змії - 300-370 мг (Д. Н. Кашкаров і Ст. Ст. Станчинський, 1940). Кількість отриманого в один прийом - сухої отрути у степової гадюки 8-10 мг, у коралової змії - 9-30 мг, у тропічних гадюк, кобр і гримучих змій - 200-300 мг і більше (Boquet, 1964). Маса сухої отрути, отриманого від Notechis scutatus,-2,1-53,9 мг (Wiener, 1960), від дорослої V. lebetina з Марокко -94 мг (Duguy, 1958), від Tr. gramineus - 1-23 мг. При електростимуляції у Cr. atrox отримано 354 мг, а без неї - 228 мг сухої отрути. Кількість виділюваного зміями отрути при укусі по-різному (С. В. Огнев, 1939; Д. Н. КаШкаров, В. о. Станчинський, 1940, та ін): у кобри 0,04-0,19 г (в середньому 0,08 м), степової гадюки - 0,005-0,03 г (0,01 г), у гюрзи -0,01-0,3 (0,06 г), у ефи -0,008 - 0,05 г (0,02 г), у щитомордника -0,003-0,1 г (0,04 г).
")