Наявність гіпоталамічного речовини, що пригнічує віддачу LH гіпофізом, визнається не всіма.
Результати експериментів з відділенням гіпоталамуса від гіпофіза показують, що гіпоталамус надає гнітюче вплив на віддачу лютеотропного гормону (LTH). Останнє пов'язується з виробленням IF - LTH або, як його інакше називають, - PIF (prolactin inhibiting factor). Наявність цього чинника підтверджується низкою експериментів. Так, наприклад, якщо лактуючих самиці ввести екстракт з медіальної эминенции, настає блок віддачі LTH. Відомо пригнічувальну дію резерпіну на гіпоталамус. Якщо ж подіяти «а тварин екстрактами гіпоталамуса інших тварин, які отримували попередньо резерпін, то в цих випадках збільшується викид LTH. Мабуть, цим пояснюється ефект, отриманий І. А. Мажбиц (1970) при лікуванні резерпіном жінок, які страждають гиполактацией.
Таким чином, можна уявити, що гіпофіз виділяє LTH, контрольований у своїх кількостях затримує гіпоталамічним фактором. Виявити активує гіпоталамічний фактор щодо LTH вдалося тільки у птахів, тому його наявність у ссавців і людини ставиться під сумнів. У той же час слід відзначити, що руйнування окремих гіпоталамічних ділянок може не супроводжуватися посиленням, а, навпаки, ослабленням продукції лютеотропного гормону (Я. М. Шинок, М. М. Нікітіна, 1962), що свідчить на користь можливого стимулюючого впливу на виділення LTH з боку гіпоталамуса.
Досить складні взаємини, що складаються між виділенням LH і LTH. Відомо, що підвищене виділення LTH призводить до зменшення LH і, навпаки, підвищення виділення LH призводить до зменшення LTH. Подібне положення може бути пояснено, якщо припустити що RF - LH, стимулюючи виділення лютеїнізуючого гормону, одночасно викликає пригнічення секреції лютеотропного.
Зіставлення різних експериментальних даних показує, що в регуляції гонадотропної функції гіпофіза можуть брати участь різні відділи гіпоталамуса, особливо передній і середній відділи; при цьому різні відділи гіпоталамуса можуть перебувати в певних субординаційних стосунках.
Участь кількох гіпоталамічних утворень у регуляції гонадотропних функцій і можливість однакового ефекту в результаті впливів, доданих до антеромедиальной зоні гіпоталамуса, змушує припускати, як це пише Б. В. Альошин (1971), що імпульси з цього відділу спочатку досягають основного центру (можливо, п. arcuatus), в якому вони інтегруються. Потім хемомедиаторный продукт передається через медіальну эминенцию в аденогіпофіз. За схемою Barraclough, Gorski (1961) комплекс, що складається з аркуатного і вентромедіального ядра гіпоталамуса, являє собою нижчий гонадотропний центр, що забезпечує базальное виділення гонадотропних гормонів гіпофіза. Але, щоб лютеїнізуючий гормон виділявся циклічно, потрібна активація зазначеного комплексу впливом другого (вищого) гіпоталамічного центру, локалізуються в преоптичної зоні (рис. 3).
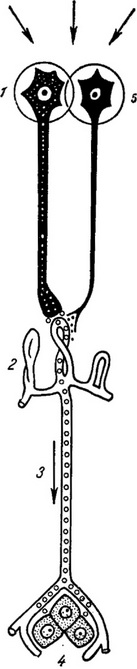 |
Рис. 3. Взаємодія нейросекреторных і нервнопроводниковых впливів у регуляції передньої частки гіпофіза (за Scharer, 1965). На виділення нейросекреторных гранул, що містять активатори гонадотропної функції аденогіпофіза, можуть впливати нервові імпульси, що надходять до медіальної эминенции з нервових клітин. 1 - нейросекреторная клітка; 2 - капілярні петлі в медіальній эминенции; 3 - портальна вена; 4 - передня частка гіпофіза; 5 - нервова клітина. |
")