Таким чином, слід зробити висновок, що регулювання гонадотропних функцій забезпечується взаємодією кількох супідрядних гіпоталамічних утворень, які можуть надавати або стимулюючий або пригнічуючий вплив як через нервові, так і гуморальні фактори. Слід вважати, що активація вищого центру щодо лютеїнізуючого гормону відбувається, мабуть, переважно нервовим шляхом.
Гіпоталамічне вплив на гіпофіз здійснюється гуморальним шляхом допомогою нейросекретов; продукуються переважно антеромедиальными зонами гіпоталамуса і скапливающимися в медіальній эминенции воронки гіпофіза.
Необхідно коротко зупинитися на проблемі можливого впливу нейрогипофизарных гормонів на продукцію аденогіпофіза. Це тим більше важливо відзначити, що при розгляді деяких клінічних моментів (див. главу 7) впливу подразнення шийки матки у жінок відзначено безсумнівний ефект у вигляді підвищення скорочувальної здатності матки. Остання ж обставина зв'язується з викидом нейрогипофизом окситоцину і вазопресину. Вазопресин, в свою чергу, потрапляючи до медіальної эминенции, діє на проникність залізистих клітин аденогіпофіза, на тонус розгалужених в ньому судин і тим самим, виступаючи як кофактор, підсилює інтенсивність дії гіпоталамічних нейросекретов (Kobayashi, 1966).
Питання про вплив внегипоталамических утворень головного мозку на функції гіпоталамуса є дуже важливим, так як гіпоталамус має численні зв'язки з корковими відділами головного мозку. Відомо, що розвиток експериментального неврозу призводить до змін эстральных циклів (Е. Б. Павлова, 1963). Подразнення електричним струмом головного мозку призводить до овуляції у кролиць (Marshall, Verney, 1951). Ін'єкція сольового розчину в кору посилює віддачу лютеїнізуючого гормону i (Taleisnik, Caligaris, De Olmos, 1962, та ін). З іншого боку, відомо, що видалення кори півкуль у щурів-самок не порушує эстральные цикли і здатність до запліднення, вагітності та пологах, хоча материнський інстинкт при цьому послаблюється (Н. Ст. Ромодановская, 1958), а вага яєчників і матки при цьому не зменшується (Greer, Schull, 1957). Подібні суперечливі дані залежать, ймовірно, від того, що безпосередньо кора головного мозку не має впливу на функцію гіпофіза з наступними змінами функції яєчників.
Дещо по-іншому позначаються впливи, виходять з більш древніх структур коркового відділу головного мозку (рис. 4). Зокрема, вплив на мигдалеподібні ядра ринэнцефалолимбической системи позначається на эстральные цикли, аж до їх припинення при видаленні ядер. Мигдалеподібний комплекс стимулює виділення як FSH, так і LH. Вимикання нюхових подразнень наркотизированием слизової оболонки носа в ряді випадків призводить до розвитку так званого назогенитального синдрому - настання помилкової вагітності, отже, до викиду значних кількостей LH. Видалення нюхових цибулин у щурів-самок призводить до атрофії яєчників (Whitten, 1956). Аналогічні реакції можна отримати, впливаючи на гіпокамп.
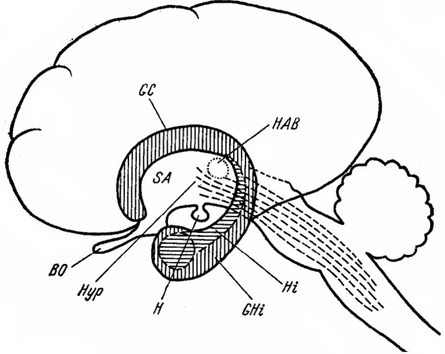
Рис. 4. Схема проекції лімбічної і ретикулярної систем вищих ссавців (Schreiber, 1963).
Лімбічна система, проекція якої заштрихована, кільцеподібно охоплює стовбур головного мозку. Пунктирною штрихуванням показана ретикулярна система, яка простягається до гіпоталамуса.
")