Запропонована тут гіпотеза може навіть пояснити результати двох улюблених демонстрацій прихильників теорії еферентної копії. Розглянемо випадок пасивного зміщення очного яблука. По-перше, я взагалі заперечую, що хто-небудь може створити пасивне рух ока, яке буде схоже на активний рух ока відносно своїх зорових ефектів. Нормальний рух ока являє собою обертання, воно не викликає зміни у відносному проекційному положенні себе і об'єктів. Як ви самі можете переконатися, пасивні рухи очей призводять до змін відносного проекційного положення спостерігача та інших об'єктів, що, згідно запропонованої нами гіпотезою, є умовою сприйняття руху. Що стосується результатів, отриманих в ситуації обмеження рухливості очних яблук, то я не можу сказати краще Вільяма Джеймса:
«Мені дуже хотілося, але сам я не зміг успішно повторити цей досвід - не знаю, з якої причини. Але навіть по відношенню до тих випадків, коли він вдавався, я повинен зауважити, що його умови занадто складні, щоб можна було з упевненістю робити ті висновки, які робить професор Мах. Замазка і тиск, який вона чинить на очне яблуко, повинні приводити до виникнення досить сильних периферичних відчуттів. Ці відчуття можуть пояснити будь-які прояви помилкового сприйняття положення очних яблук абсолютно незалежно від почуття іннервації, існування якого припускає професор Мах» (Джеймс, 1890) [7].
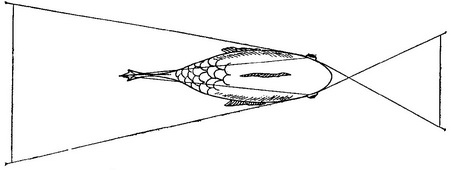
Психологи схильні думати про «Я» або як про безглуздою абстракції, або як про складну проблему, що вимагає глибокого роздуму. Зведення «Я» до зорової проекції носа на сітківку зазвичай викликає радісне пожвавлення і глузування слухачів. Навіть Гібсон (1950), який зробив для розуміння ролі проекції носа більше, ніж який-небудь інший психолог, писав про це зі свого роду захисною обережністю. Я хотів би сказати тут кілька слів на захист носа. Серед різних видів тварин з камерними очима немає жодного, чиї очі не отримували б проекцію носа або який-небудь іншої частини тіла. У всіх представників видів з розвиненим панорамним зором панорама переривається проекцією самої тварини. У представників хижих видів, відрізняються максимальною фронтальностью положення очей і максимальним бінокулярний перекриттям, завжди є проекція себе. Це не може бути просто біологічної випадковістю. Проекція частин тіла на сітківку повинна мати важливе функціональне значення, інакше вона б не збереглася у багатьох еволюційно різних форм. У самому справі, запропонована тут гіпотеза про сприйнятті положення має те гідність, що вона надає сенс фактом, який інакше здається тривалої біологічної помилкою (Уолс, 1942) (див. рис. 3.18) [8].
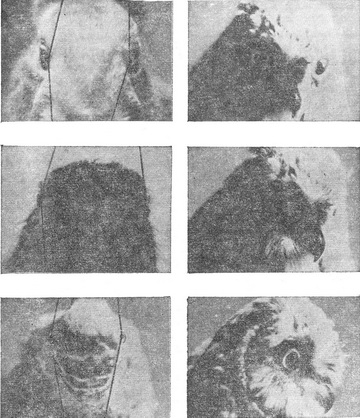 |
Рис. 3.18. Зорове поле різних тварин. У зайця (зліва вгорі і в середині) і вальдшнепа (зліва внизу), які є жертвами для багатьох інших тварин, розвинулося виключно широке зорове нулі, що охоплює до 200°. Внаслідок цього, як заєць, так і вальдшнеп можуть одночасно бачити свій ніс (дзьоб) і хвіст. Риба має загальне зорове поле близько 360°. Значна його частина зайнята тілом і плавниками риби. Сова має зоровий кут 110° с бінокулярний перекриттям, її очі спрямовані прямо вперед. Однак у цієї птиці є спеціальний назальний чубчик, який займає центральні 30° її зорового поля. |
Проксимальна інформація, яку ми описували досі, зрозуміло, являє собою не єдиний вид інформації, який може специфікувати положення прямо перед головою. Гібсон (1950) описав стимульні наслідки руху спостерігача у напрямку до об'єкта і стимульні наслідки руху об'єкта до спостерігача. Розглянемо ситуацію, в якій спостерігач дивиться на текстуровану поверхню стіни, подібну до показаної на рис. 3.19. Як це видно з малюнка, коли спостерігач рухається у напрямку до стіни, проекційне зображення на його сітківці буде систематично змінюватися.
")